

中国农业科学院博士后揭示大莎草C3–C4光合作用转换遗传基础和莎草科核型演化
与C3植物相比,C4植物具有更高的光合效率和氮素、水分利用率。将C4途径导入C3作物,预计可以大幅提高其光合效率和产量 (Hibberd et al., 2008)。然而,C3和C4植物在叶片结构和生化途径等多个方面存在明显差异,目前有关C3向C4进化和转变的遗传机制仍不清楚。大莎草 (Eleocharis vivipara) 是一种莎草科无叶、两栖植物,其具有多种显著特征,最显著特征是在水下使用C3光合类型,而在陆地使用C4类型 (Chen et al., 2011; Ueno et al., 1988),但目前因缺乏基因组数据而阻碍了其光合作用转换的遗传研究。
近日,JIPB在线发表了中国农业科学院深圳农业基因组研究所程时锋团队题为“The genome of Eleocharis vivipara elucidates the genetics of C3–C4 photosynthetic plasticity and karyotype evolution in the Cyperaceae”的研究论文 (https://doi.org/10.1111/jipb.13765),该研究首次完成了大莎草染色体级别基因组组装和注释,并通过比较和进化分析等策略为C3–C4光合作用转换机制提供了重要见解 (图1)。
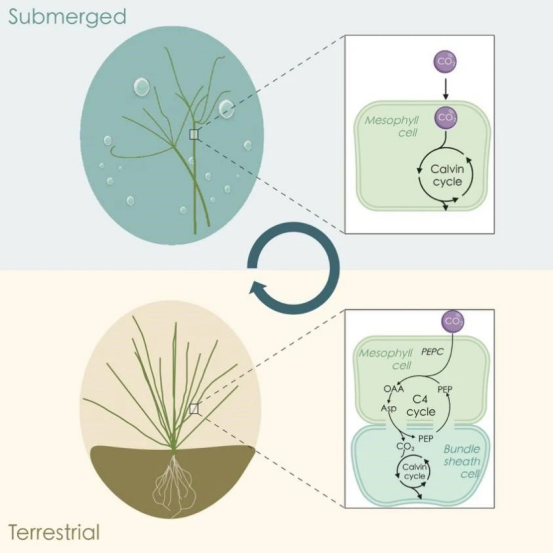
图1 大莎草水生C3和陆生C4光合作用转换模式图
研究发现,大莎草基因组大小约为965.22 Mb,有38,769个蛋白编码基因,是一个四倍体,染色体数目为2n = 4x = 20 (图2)。在~3,296万年前,大莎草与近缘物种油莎草 (Cyperus esculentus) 分化后,又在~350万年前经历了一次全基因组复制事件。与大莎草相比,其多个近缘物种经历了频繁的染色体融合和断裂事件。而大莎草与其近缘物种的全着丝粒,则有利于维持其重排后的染色体稳定性,并促进物种形成,从而在一定程度上解释了莎草科物种数量 (~5,400种)、染色体数目 (n = 2–113) 和倍性丰富的原因。
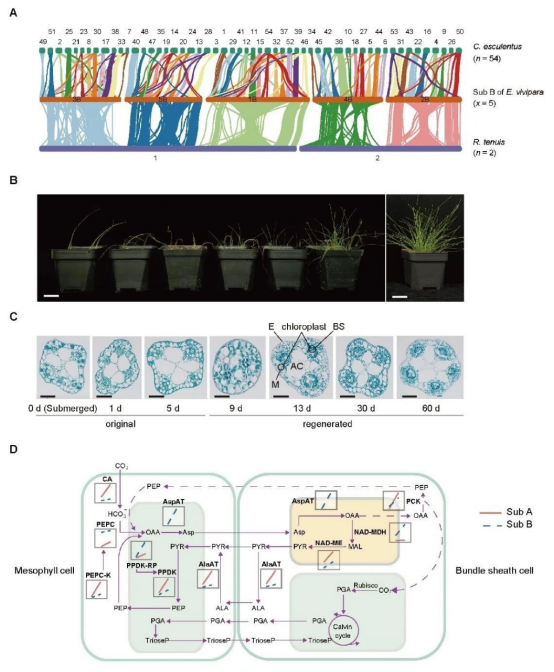
图2 莎草科核型演化和大莎草C3–C4光合可塑性分子机制
将大莎草从水生移栽到陆生实验显示,其茎秆经历了干枯、再生和稳健生长发育过程 (图2)。再生过程中,茎秆能够形成花环结构这一C4光合典型特征。基因共表达调控网络分析发现,在陆生茎秆中,C4核心通路基因总体表达增强,
与可能驱动C4进化或转变的光呼吸、缺水和脱落酸响应等相关基因,以及与光合效率提高的花环结构改良、叶绿素合成、光捕获和碳固定等基因显著富集,表明这些基因在由C3转换至C4光合过程中发挥了潜在重要作用。研究还发现,大莎草中存在混合型NAD-ME和PCK C4亚型,其可在多种光照条件下维持较高光合效率,且该亚型由A亚基因组主导和B亚基因组辅助。此外,系统进化结果显示,虽然其C4光合作用出现早于全基因组复制事件,但复制后显著扩张基因有利于提高其光捕获和光合效率等能力。
大莎草的这种光合可塑性能力,与仅使用C3类型的水稻和C4类型的玉米等其他物种形成鲜明对比。除大莎草外,这种C3和C4光合作用在单个个体的不同器官或生命周期的不同阶段同时发生事件,目前仅发现于禾本科和藜科内的少数其他植物 (Keeley, 1998; Pyankov et al., 2000)。这种在单个基因组背景下的光合作用转换和不同基因表达机制,表明该复杂性状可能受到表观遗传等因素调控。本研究不仅为此类可塑型光合作用植物提供了首个高质量参考基因组,为富含C3–C4中间表型材料 (如黄顶菊和百簕花等) 提供了宝贵补充,更为深入理解C3向C4光合作用转变的遗传机制提供了重要线索,为未来作物改良和育种工作提供了新的思路。
中国农业科学院深圳基因组研究所博士后刘红兵、博士生赵航、硕士生张燕文、科研助理黎秀丽和中国科学院植物研究所助理研究员左毅为论文共同第一作者,中国农业科学院深圳基因组研究所程时锋研究员与美国密歇根州立大学博士后Matt Stata为共同通讯作者。中国农业科学院生物技术研究所路铁刚研究员、中国科学院植物研究所王雷研究员和加拿大多伦多大学Rowan Sage教授对该研究提供了重要指导。本研究得到国家重点研发计划和国家自然科学基金等项目的资助。